發表期刊:Nature Communications
影響因子:12。353
發表時間:2019年
合作單位:第三軍醫大學附屬西南醫院
今天百趣代謝組學將給大家分享Nature Communications上的一篇文章:ABHD5 blunts the sensitivity of colorectal cancer to fluorouracil via promoting autophagic uracil yield。
百趣代謝組學分享,氟尿嘧啶(Fu)是最常用的治療直腸癌(CRC)的藥物。然而因藥物抗性,極大限制了其治療的療效。自噬(macroautophagy)是一種細胞分解代謝過程,自噬有潛力為附近所有代謝途徑供能,為細胞提供巨大的代謝可塑性。代謝重程式設計和代謝酶的異常活躍被認為是惡性腫瘤的特徵之一。
作者證明ABHD5定位於溶酶體後與PDIA5相互作用,防止PDIA5與RNASET2相互作用,從而使RNASET2失活。代謝組學分享,它會損害RNASET2介導的自噬尿嘧啶產量,促進結直腸癌細胞將FU作為外源尿嘧啶攝入,從而增加其對FU的敏感性。研究結果首次揭示了ABHD5在調節溶酶體功能中的新作用,對基於FU輔助化療後的機體恢復提供了重要見解。
ABHD5降低了CRC細胞對FU的敏感性

圖1 ABHD5降低了CRC細胞對FU的敏感性
採用癌症藥物敏感性基因組學資料集,將ABHD5表達水平與化療結果相關的敏感性資料集集合起來,透過Pearson’s correlations的分析方法,ABHD5表達在pMMR和dMMR CRC細胞系中都和IC50呈現正相關(圖1a)。代謝組學分享,相比較與正常的癌細胞,ABHD5基因敲除的細胞(ABHD5 KD)在FU處理下IC50,細胞活力顯著降低(圖1b-c),細胞凋亡率顯著升高(圖1d)。
隨後作者做了小鼠實驗,透過生物發光成像和TUNEL染色,得到同樣的結論,ABHD5 KD組小鼠對FU處理後腫瘤負荷明顯減小(圖e),並且組織中腫瘤細胞凋亡率增加(圖f)。代謝組學分享,對不同的小鼠做ABHD5基因敲入和敲除後,用PBS或FU處理,每隔一段時間給小鼠和腫瘤組織稱重,發現ABHD5 KD+FU處理組明顯病情得到緩解(圖1g-h)。再透過Caspase-3對凋亡細胞或Ki67對增殖細胞做免疫組化檢測,也得到相同的結論ABHM5 KD+FU處理明顯抑制了細胞的增值並表現較強的敏感性(圖1i-j)。
FU輔助的化療效果受ABHD5表達量影響

圖2 ABHD5的表達以及FU輔助化療的療效
將NCBI-GEO資料集中361名CRCs患者分為ABHD5 high組(262例)和ABHD5 low組(99例),統計結果顯示ABHD5 high可能是BRAF陽性突變(BRAF MT),p53野生型(p53 WT)和染色體不穩定陰性型(CIN-)(圖2a-b)。只用FU治療的ABHD5 high效果較好,有化療的還是ABHD5 low組更好(圖2c-d)。代謝組學分享,用另外一組大資料量進行驗證,也是同樣的結果,ABHD5 low組更適合FU+化療(圖e-f)。
ABHD5透過提高自噬性
尿嘧啶的產量來控制FU的攝取

圖3 ABHD5影響CRC細胞對FU的攝取
透過HPLC發現ABHD5 KD細胞中FU濃度增加,WB顯示PBS或者FU處理的正常癌症細胞中都沒有發現TS, TP,DPD(這3個酶控制FU代謝)的變化,可能是FU濃度和藥物吸收能力相關(圖3a-b)。代謝組學分享,但是FU處理後,ABHD5 KD細胞尿嘧啶減少(圖3c),但是尿嘧啶生物合成限速酶CPS II和UMPS沒有發生變化(圖3d)。ABHD5可能透過其他的方式影響了尿嘧啶的產量。
在GSE38832資料集122例CRCs,ABHD5 low組溶酶體途徑相關基因是下降的(圖3e),根據資料做了個熱圖,猜想溶酶體RNA的降解使得細胞產生了自噬,從而提高了尿嘧啶的含量(圖3f)。HCS分析顯示自噬體的量和FU的含量呈現負相關趨勢(圖3g-h)。代謝組學分享,ABHD5過表達的細胞在FU和靶向藥物處理後,才使得胞內尿嘧啶和FU的濃度提高(圖3i-j)。在PDX模型中BECN1啟用劑對ABHD5 KD中的尿嘧啶和FU的濃度影響不是很大(圖3k-l),說明ABHD5透過其他途徑調節細胞自噬。
LC-MS檢測ABHD5透過
RNASET2來促進細胞自噬

圖4 RNASET2調控ABHD誘導細胞自噬
採用LC-MS檢測FU作用下尿嘧啶和相關的核苷含量的變化,ABHD5 KD組中核苷和尿嘧啶的含量沒有什麼變化,可能是RNASET2催化RNA分解過程並參與ABHD誘導細胞自噬(圖4a-c)。代謝組學分享,ABHD5 KD和RNASET2 KD組中核苷產生後一會就沒有了,ABHD5高表達+RNASET2 KD組中反而對FU敏感了(圖4d-g)。這些結果表明,ABHD5透過RNASET2來促進細胞自噬,使得尿嘧啶含量增高。
ABHD5保護RNASET2不被PDIA5滅活

圖5 ABHD5維持RNASET2的活性
透過免疫熒光染色和WB檢測說明ABHD5定位於溶酶體並調節RNASET2的活性(圖5a-b)。代謝組學分享,酵母雙雜實驗表明ABHD5不直接和RNASET2直接作用,而且ABHD5 KD的細胞中RNASET2表達量沒有變化(圖5c-e)。透過蛋白晶片檢測,表明ABHD5與RNASET2是共同競爭結合PDIA5的關係,從而促進細胞自噬,減弱CRC細胞對FU的敏感性(圖5c-m)。
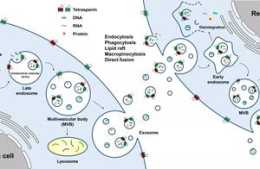
結論

圖6 文章機制示意圖
ABHD5定位於溶酶體,並且和RNASET2共享PDIA5中的相同相互作用域。 ABHD5直接與PDIA5相互作用,以防止PDIA5直接與RNASET2相互作用並使RNASET2失活。ABHD5缺乏釋放使得PDIA5直接與RNASET2相互作用,使RNASET2處於失活狀態,這會損害RNASET2誘導的自噬尿嘧啶產量。 ABHD5缺乏促進CRC細胞攝取FU作為外源性尿嘧啶,從而增加其對FU的敏感性。 代謝組學分享,相反,由於自噬尿嘧啶產量增加,ABHD5使得的CRC細胞顯示出對FU的抗性。
LC-MS非靶標代謝組學檢測
BIOTREE LC-MS非靶標代謝組學檢測透過液質聯用(LC-MS)方法檢測生物體受外界刺激前後體內大多數小分子代謝物的動態變化,重點尋找在實驗組和對照組中有顯著變化的代謝物,進而研究這些代謝物與生理病理變化的相關關係,其研究物件大都是分子量1500Da以內的小分子物質。